Out of over 3 trillion trees globally, more than 40% occur in (sub)tropical settings that contribute to two-thirds of all natural CH4 emissions.

The role of soil and non-woody plants in transporting methane (CH₄) axially from rhizosphere to the atmosphere as a “passive pipe” is well understood and represented in global carbon budgets and Earth System Models (ESMs). On the other hand, CH4 transport through woody plants is highly uncertain and underrepresented in ESMs. In the past few years, some important studies, particularly published in New Phytologist, have enriched our understanding of CH4 dynamics within the caulosphere (i.e., tree bark, Figure 1). Radial diffusion of CH4 from tree stems is acknowledged as "a new frontier in the global carbon cycle" and is sometimes referred to as treethane.
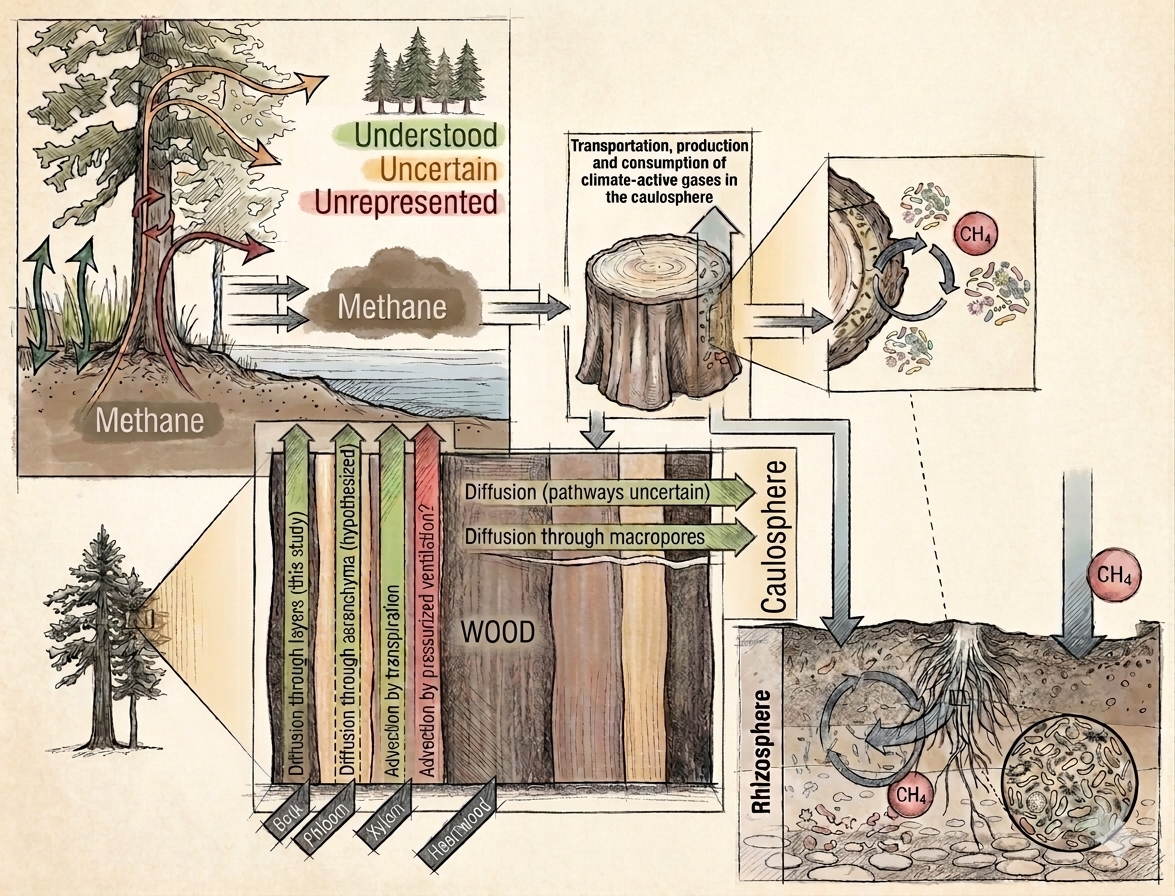
Figure 1: Methane dynamics at the caulosphere-rhizosphere interface in woody vegetation (adopted from Ward and Megonigal, 2024; Leung et al., 2026)
Despite spanning a global surface area approximating 143 (±59) million km2, close to the total land surface area of 149 million km2, the caulosphere is an understudied component of soil-tree-atmosphere interactions. Out of over 3 trillion trees globally, more than 40% occur in (sub)tropical settings that contribute to two-thirds of all natural CH4 emissions. Accordingly, there is growing consensus that methane dynamics in woody vegetation must be evaluated and considered in ESMs, along with process-based models.
Tree Stems as Sources of CH4
Further investigation into methanogenesis (i.e., CH4 production) within the caulosphere has revealed that trees harbour active and diverse microbial communities capable of metabolizing a range of climate-active gases. Notably, fungal-mediated heartwood decay has been identified as a key driver of CH4 emissions from living trees. Fungal CH4 production is linked to chloromethane (CH3Cl). Some submergent macrophytes such as mangroves are known to thrive under anoxic soil conditions and therefore act as CH4 sources. Anoxia may be created through oxygen-consuming metabolic processes of tree-associated endophytic microbes, such as fungi, as well as through active stem and root respiration. Another recently discovered CH4-producing group is cyanobacteria, which have been linked to this process both under oxic and anoxic conditions, depending on substrate availability.
Tree Stem as Sinks of CH4
While on one hand, tree stems emit substantial CH4 to the atmosphere, there exists another mechanism, regulated by bark-dwelling microbes of lowland trees, that oxidizes CH4. Though the significance of methane oxidizing bacteria (MOB) in soil and water is widely known, trees are a lesser-known reservoir, not because of the general absence of these populations, but at least partly due to underdeveloped detection methods with poor sensitivity. Acknowledging the seminal work of Luke Jeffrey and others on the discovery of MOB in Melaleuca quinquenervia (paperbark tree, native to Australia), George A. Inglish described this finding as “bacteria in tree bark are hungry for methane.” These methanotrophic bacteria can reduce up to one-third of total treethane emissions. According to the most recent meta-analysis, the global mean woody surface CH4 sink is estimated at 37.2 Tg of CH4 yr−1 comparable to the soil sink (30–40 Tg of CH4 yr−1, 1 Tg = 1012 g).
Since the concept of tree-stem CH4 dynamics is still emerging globally and lacks clear empirical validation in the Indian context, a structured Q&A-based knowledge-sharing framework could help generate more insights and support its development.
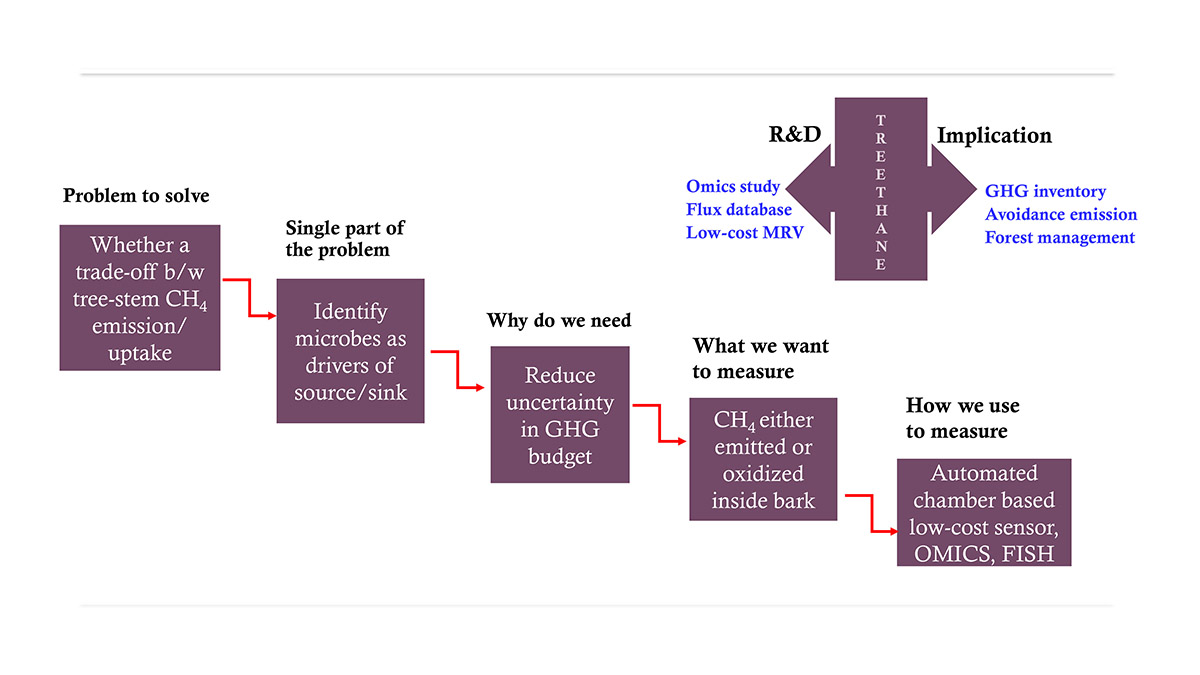
Figure 2: Conceptual framework from problem identification to real-world application.
(1) Is there a standard height of the stem from the soil surface for chamber deployment?
Generally, measurements are taken at about 2 m above ground level as a standard height. However, even at this level, chamber placement may overestimate methane emissions, especially in submerged vegetation like mangroves, because the lower parts of tree stems are closer to methane-rich soils. This methane can be taken up by roots and transported through the tree before being released into the air. Therefore, deployment must account for the tidal prism, particularly in the case of mangroves.
(2) What is the contribution of tree-stem CH4 to total emissions across forested biomes in India?
The answer is not yet well constrained, despite India hosting a diverse array of both highland and lowland floral ecosystems, including a wide range of mangroves (~0.5 million ha), cottonwood, and paperbark trees. While India encompasses a wide range of vegetated biomes, the role of tree-mediated CH4 flux remains underexplored. Recent papers published in leading journals such as Nature and Science, highlight the global relevance of tree-stem CH₄ emissions and underscore the need for region-specific investigations. These developments should encourage climate scientists and plant biologists in India to undertake long-term monitoring of tree-stem methane fluxes and integrate such data into national trace gas inventories.
(3) What would be the next steps for measurement?
Omics and data science: Promoting metagenomics and meta-transcriptomics to discern selective amplification and detection of CH4 oxidizing gene in tree stems/barks (functionally detected through extensive searches of public metagenomic libraries in India, such as National Centre for Biotechnology Information-Sequence Read Archive or NCBI-SRA).
Method development: Moving beyond manual static chambers to Automated Tree Enclosure Systems paired with low to medium cost, high accuracy sensors will enable high-frequency data collection and strengthen measurement, reporting, and verification (MRV) tools. This is crucial for capturing temporal (diurnal or seasonal) pulses of CH4 that manual sampling often misses. Omics data could be utilized to design probes for applications such as fluorescence in situ hybridization (FISH) to visualize the precise spatial distribution of CH4-cycling microbes within different tree tissues.
(4) How can this research contribute to carbon finance in the long run?
- Despite being a scientifically proven phenomenon, treethane is not integrated into international carbon registries due to a lack of standardized protocols. By identifying and sustainably planting tree species rich in MOB through rigorous research, an “avoided emissions” pathway could be established, and MRV tools such as FISH could provide direct technical validation to unlock carbon finance for these specialized ecosystems.
- By promoting carbon sequestration pathways, additionality can be demonstrated through activities such as secondary plantation of suitable tree species or the conservation and restoration of degraded lands, provided these interventions lead to measurable carbon gains beyond a defined baseline scenario.
References
- Gauchi, et al., Nature, 631, 796-802
- Jeffrey, et al., 2023. New Phytologist, 242, 49-60
- Ward, N.D and Megonigal, J.P. 2024, New Phytologist, 5-7
- Leung, P.M., 2026. Science, 8, 1-14.
- Barba, J., et al., (2019). New Phytologist, 222: 18-28
- Inglis, G.A.S. 2021. Commun Biol4, 738.